"It
has not escaped our notice that the specific
pairing we have postulated immediately
suggests a possible copying mechanism for the
genetic material."
--
J.
Watson & F. Crick, 1953
|
- What Must the Genetic Material Be Able To
Do? As we will see over the next few units,
the base pairing proposed by Watson and Crick turned
out to be very important in the work of DNA as the
genetic material. Just what is that work? What must
the genetic material be able to do? There are at
least three things.
- It Must Contain Information: DNA
must have somewhere in its molecular structure the
information that specifies what kind of organism
its bearer becomes. This is the information that
directs cellular and organismal processes. It must
have the ability to determine that you are a human
and to determine your skin pigmentation, your hair
color, and your susceptibility or resistance to
various diseases (among countless other traits).
From the time that scientists conceded that DNA
was the genetic material, it was their working
hypothesis (which turned out to be true) that this
information was in the sequence of DNA's nitrogen
bases.
- It Must Be Capable of Replicating Itself:
The information stored in a DNA molecule must be
copyable. A DNA molecule must be capable of
replicating, thus producing two molecules with an
identical base sequence. That is the topic of this
unit.
- It Must Be Capable of Determining Traits:
The information coded in the form of a base
sequence must have meaning that can be decoded.
The sequence of bases must somehow be capable of
directing the cellular activities within the
organism. That is, it must be capable of
determining what kind of cell, tissue, organ, and
organisms develops. It must be capable of
determining traits. We will see that this process
occurs by DNA directing which proteins (including
the all-important enzymes) the cell makes.
- It Must Do Some Other Things: The
three roles of the genetic material above are
absolutely essential, but actually it also must be
capable of other functions, such as undergoing
chemical and physical changes (mutation) and break
apart and joining with other DNA molecules, as
occurs during crossing over (recombination). We
will also take up these topics later.
|
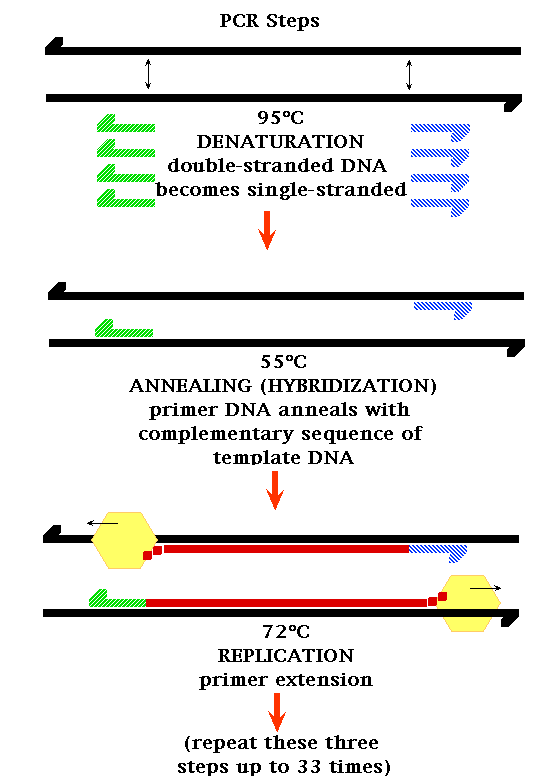
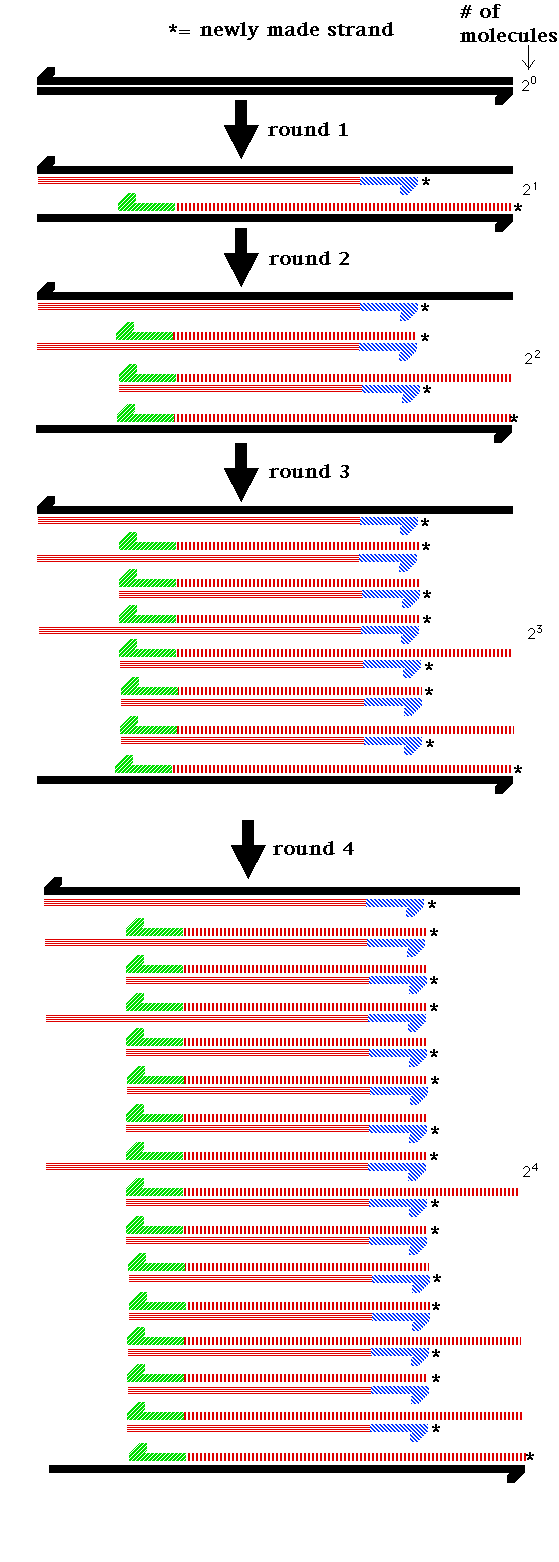
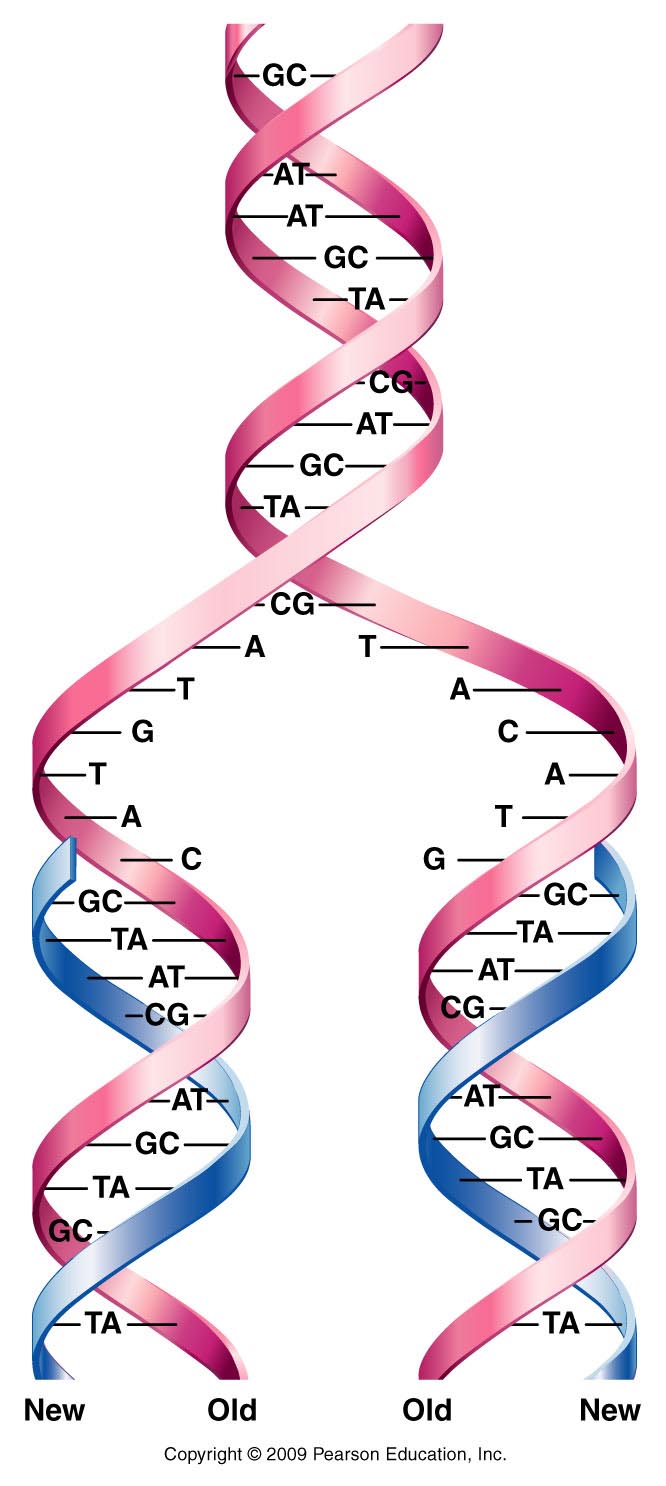
- DNA Replication: Watson and Crick
proposed a simple model of DNA replication whereby
the hydrogen bonds holding the two strands of a DNA
molecule are broken and the two strands come apart
(unwind). (This process is called denaturation and
will occur when DNA is heated to about 95ºC. The
opposite process is annealing or hybridization and
occurs when denatured DNA is slowly cooled.) Each
single-stranded molecule can then serve as the
template for the synthesis of a new strand. The new
strand has a base sequence that is complementary to
that of the template (following A-T, G-C specific
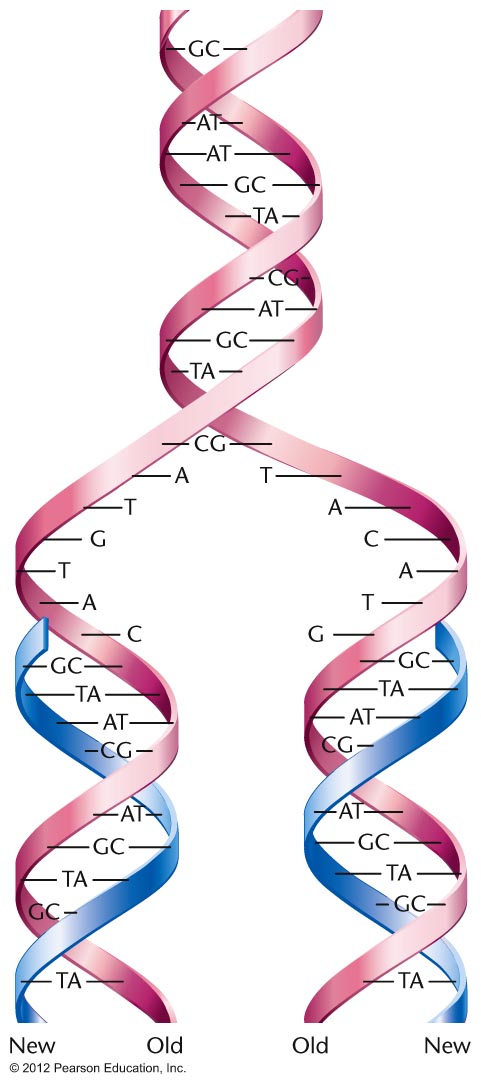
base pairing). This model of replication is the
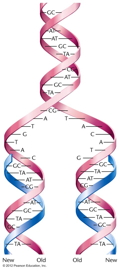
semiconservative model of DNA replication. (Each
"new" DNA double helix is actually composed of one
newly-made strand and one "old" strand. That is, the
original molecule is half conserved in the new
molecule.)
|
 |
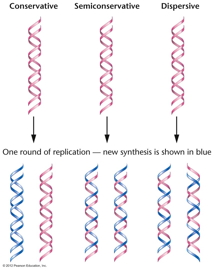
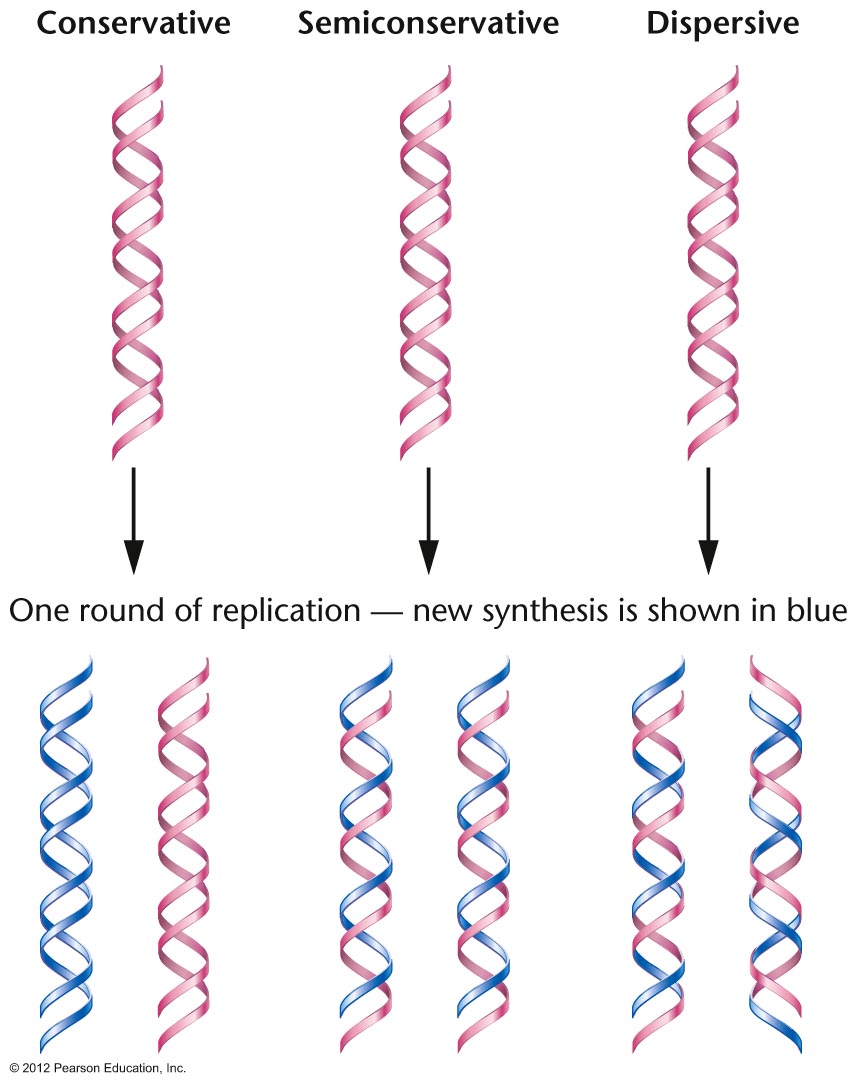
- The Proof of Semiconservative DNA
Replication: If DNA replication were not
semiconservative, the other main possibility was
that it was conservative, where an entirely
newly-made double helix is made from the original
helix. (A dispersive model was also proposed.) The
proof that Watson and Crick's idea was right first
came in studying E. coli DNA replication.
|
 |
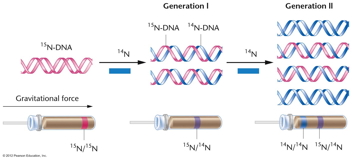
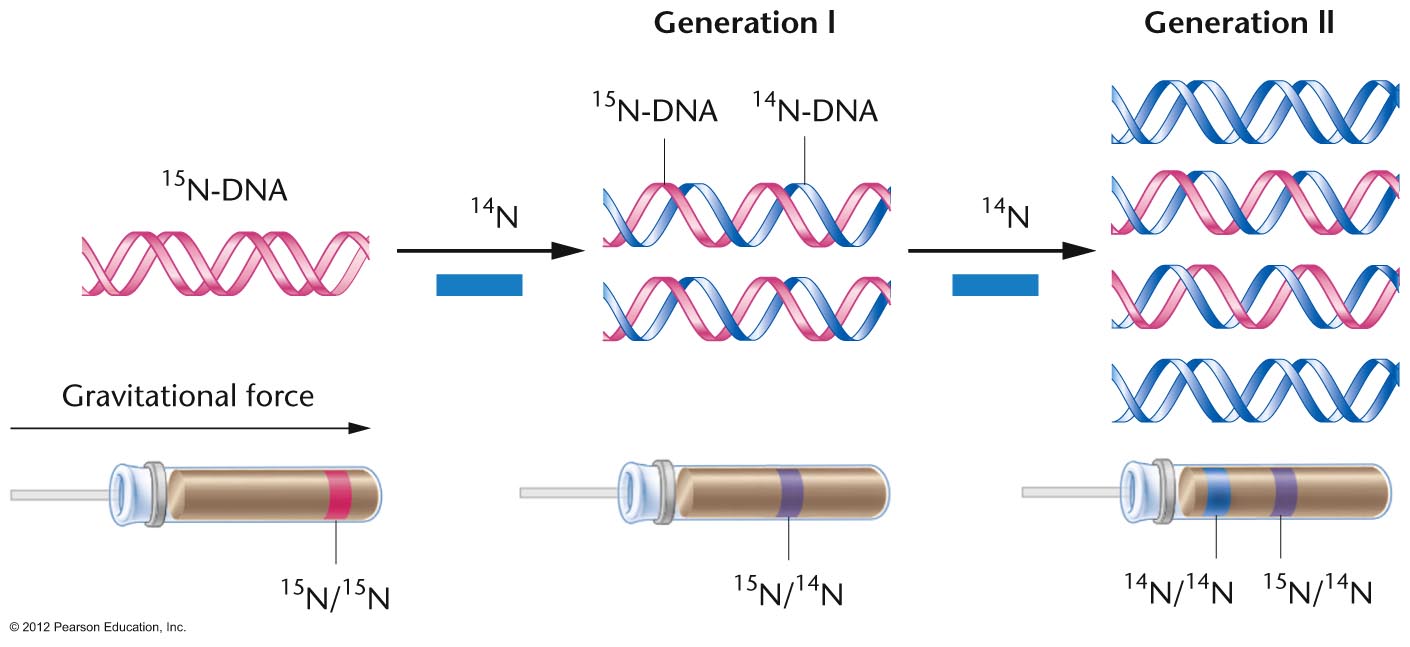
- Meselson and Stahl's Experiment in a
Prokaryote: In 1958, Meselson and Stahl
performed an experiment with E. coli
replication showing it to be semiconservative.
|
 |
- Taylor, Woods, and Hughes' Experiment
in a Eukaryote: Taylor, Woods, and
Hughes showed that DNA replication is
semiconservative in cells of the fava bean plant
using the technique of autoradiography.
|
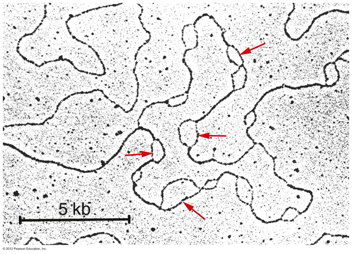
- The Origin of Replication and the
Replication Fork: DNA replication occurs
at a branch point called a replication fork.
Replication begins at a site called the
replication origin, which involves specific DNA
sequences in both prokaryotes and eukaryotes.
|
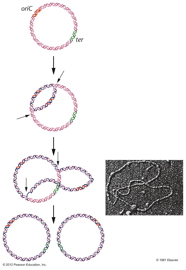
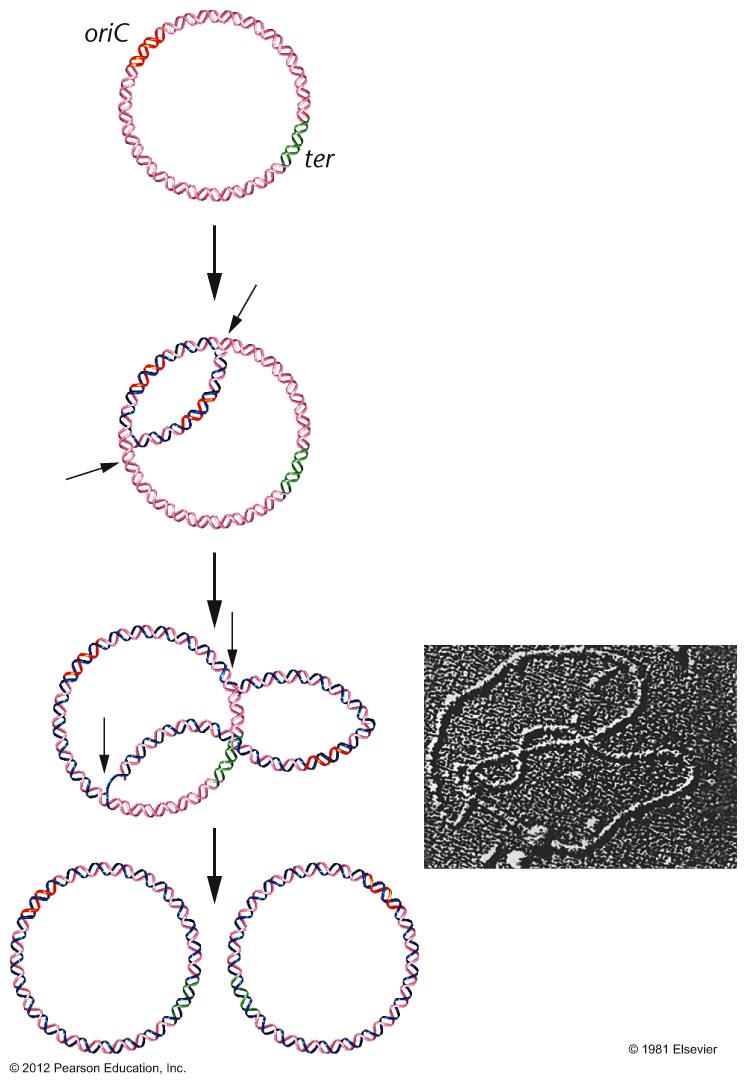
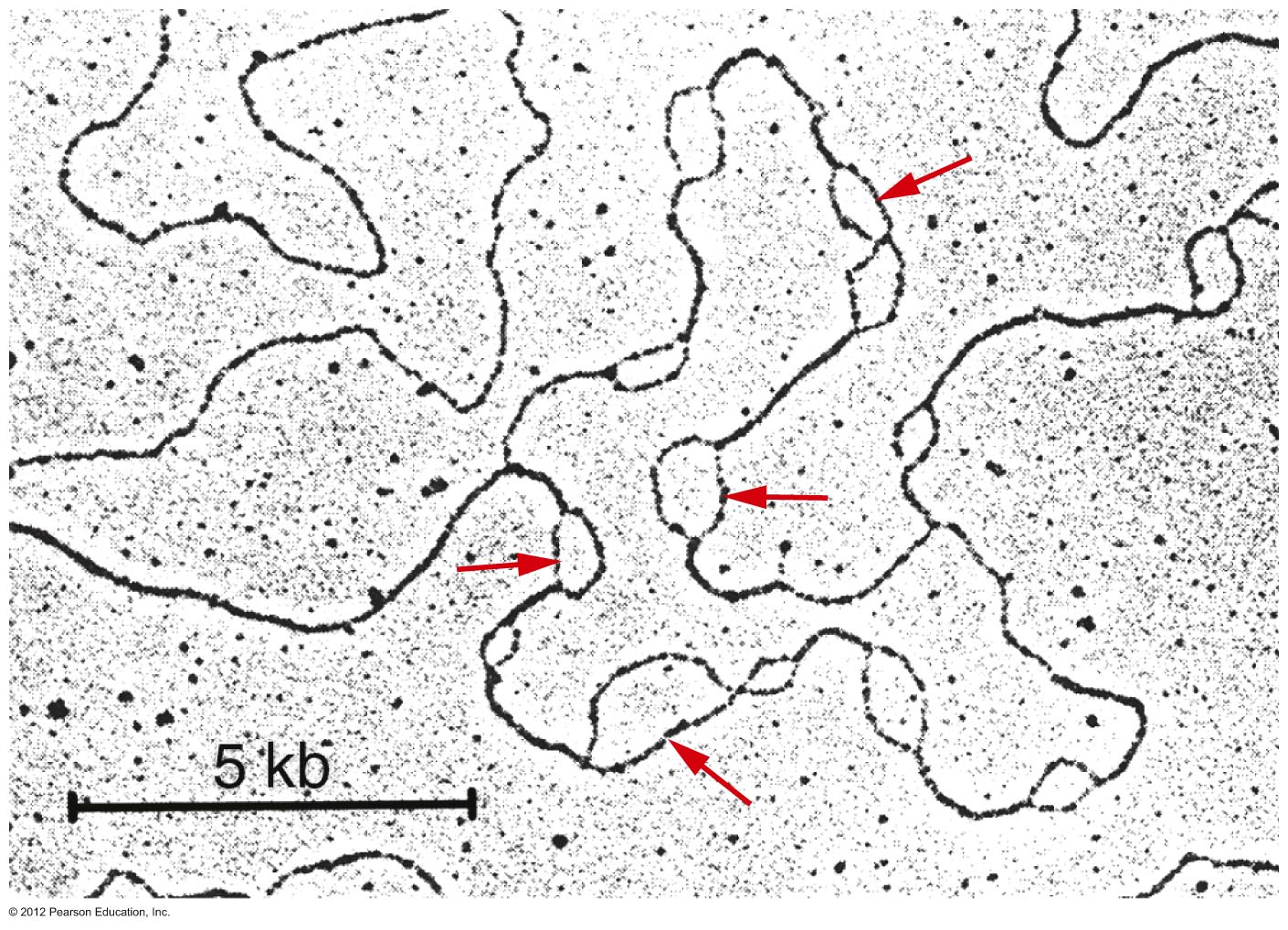
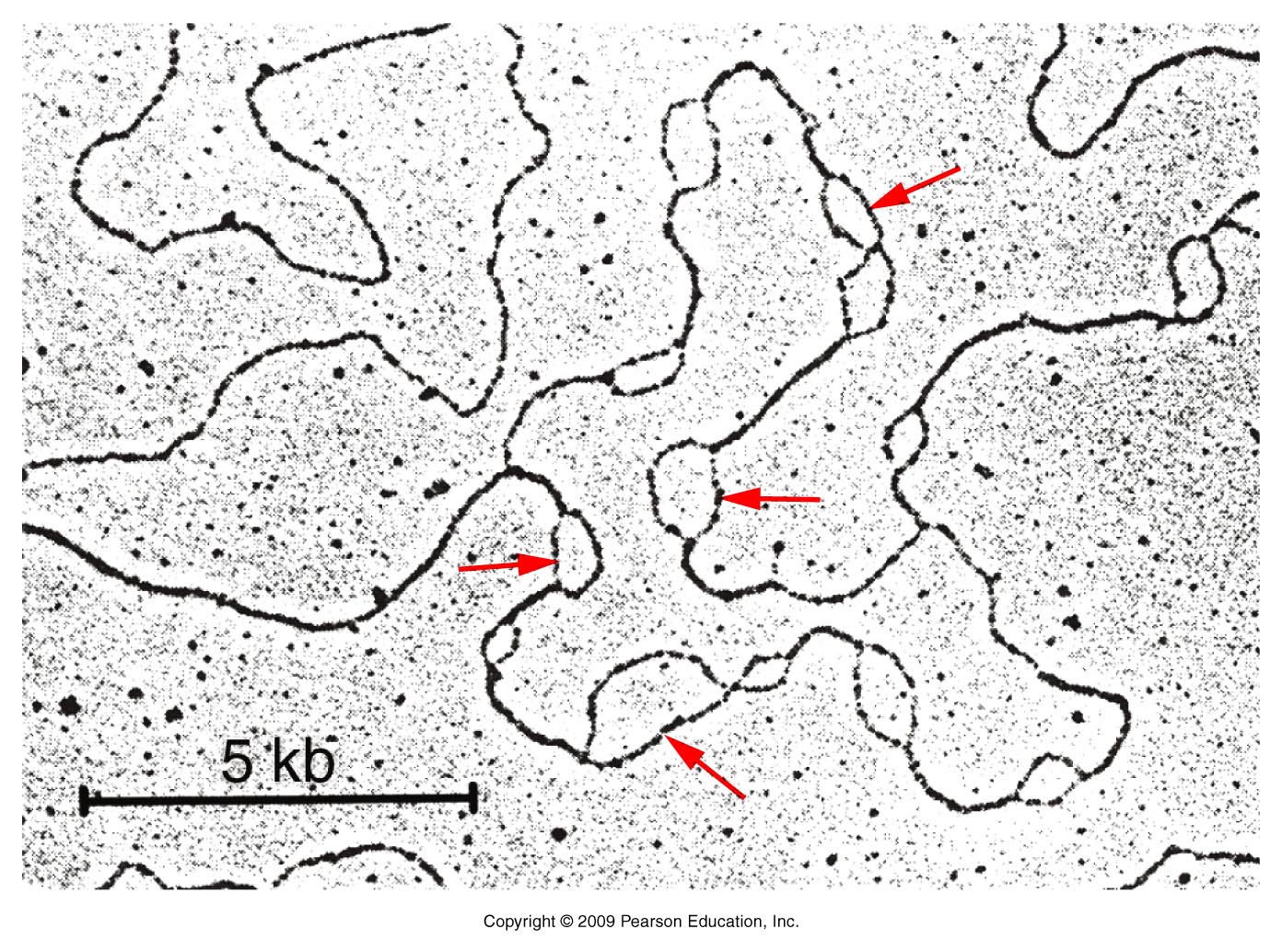
- Prokaryotes: Cairns demonstrated
that E. coli's chromosome has one
replication origin. That is, the entire circular
chromosome is one replicon. Replication from
that origin proceeds bidirectionally until the
two replication forks meet on the opposite side
of the chromosome.
|
 |
- Eukaryotes: Eukaryotic
replication begins at numerous replication
origins with mammals having about 25,000 origins
across the genome. Replication is bidirectional
from each origin.
|
 |
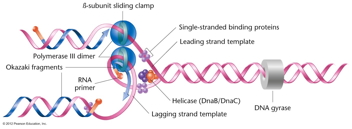
- The Enzymes of DNA Replication:
There are a number of enzymes that are required
for replication of DNA in vivo. (Video
showing how all of the enzymes below may work
together to replicate DNA)
|
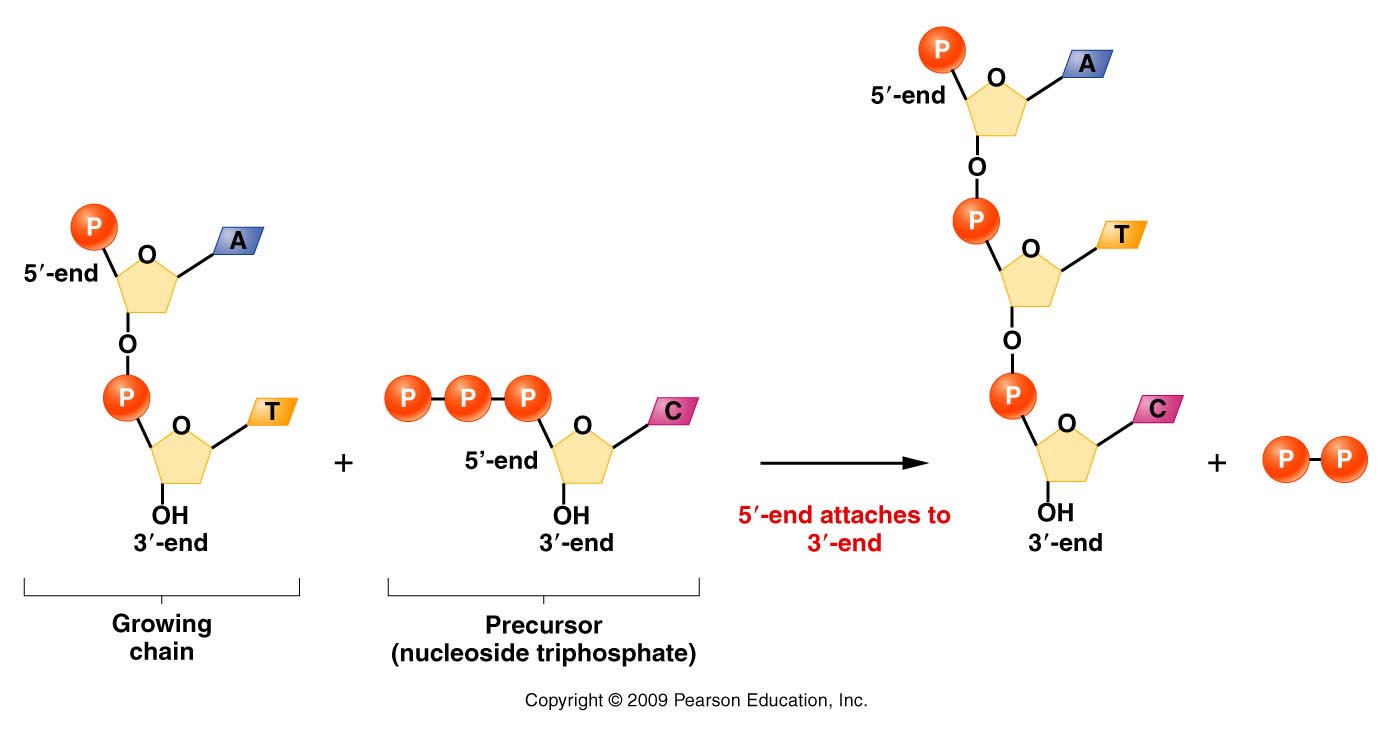
- The DNA
Polymerases: The main
class of enzymes needed for DNA replication are
the DNA polymerases. These enzymes can add a
nucleotide to the 3' OH of a primer as directed
by the template strand (base pairing). They use
the nucleoside triphosphates (dATP, dGTP, dCTP,
dTTP) with pyrophosphate being cleaved off
during the synthesis reaction. A phosphodiester
bond is formed and the 3' end of the primer is
extended (primer extension).
- Prokaryotes: The
first DNA polymerase discovered was by Arthur
Kornberg (1957) in E. coli and became known as
DNA
polymerase I. Since DNA polymerase I
mutants could still grow and replicate their
DNA, it was realized that this must not be the
main replication enzyme (it is primarily a
repair enzyme). Subsequently, DNA
polymerase II and III were discovered
(and more). DNA polymerase III turns out to
be the main in vivo replication enzyme.
- Eukaryotes:
Eukaryotes have a number of DNA polymerases
with polymerase δ
primarily responsible for in vivo
nuclear DNA replication.
|
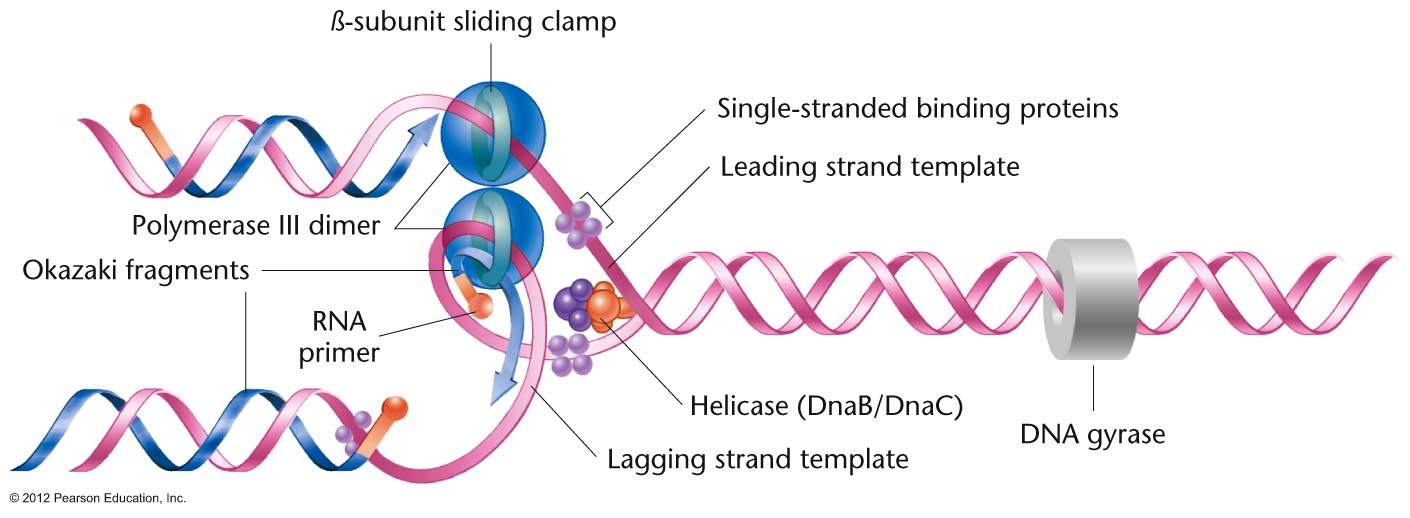
- DNA
Helicase:
This enzyme works at the replication fork to
break the hydrogen bonds and thus unwind the
DNA.
- SSBs: Since
single-stranded DNA collapses on itself,
proteins called single-strand binding proteins
bind to the denatured single strands maintaining
the strand in linear form.
- A Problem (Solved by Topoisomerases
like Gyrase): The unwinding of the
helix creates a problem, since unwinding
requires the rotation of the double helix. The
enzyme called topoisomerases solves this problem
by allowing rotation. (Topoisomerase
video)
- Another Problem
(Solved by the Enzyme Primase): The
fact
that
DNA synthesis only occurs in the 5' to 3'
direction creates another problem. Replication
of one of the strands can proceed without
problem, but the antiparallel nature of DNA
means that replication must proceed on the
other strand in the opposite direction. However,
on that strand, there is no primer to attach a
nucleotide to (which is the only thing DNA
polymerase can do).
|
|
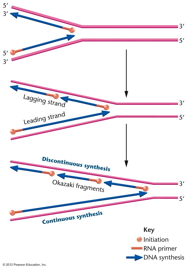
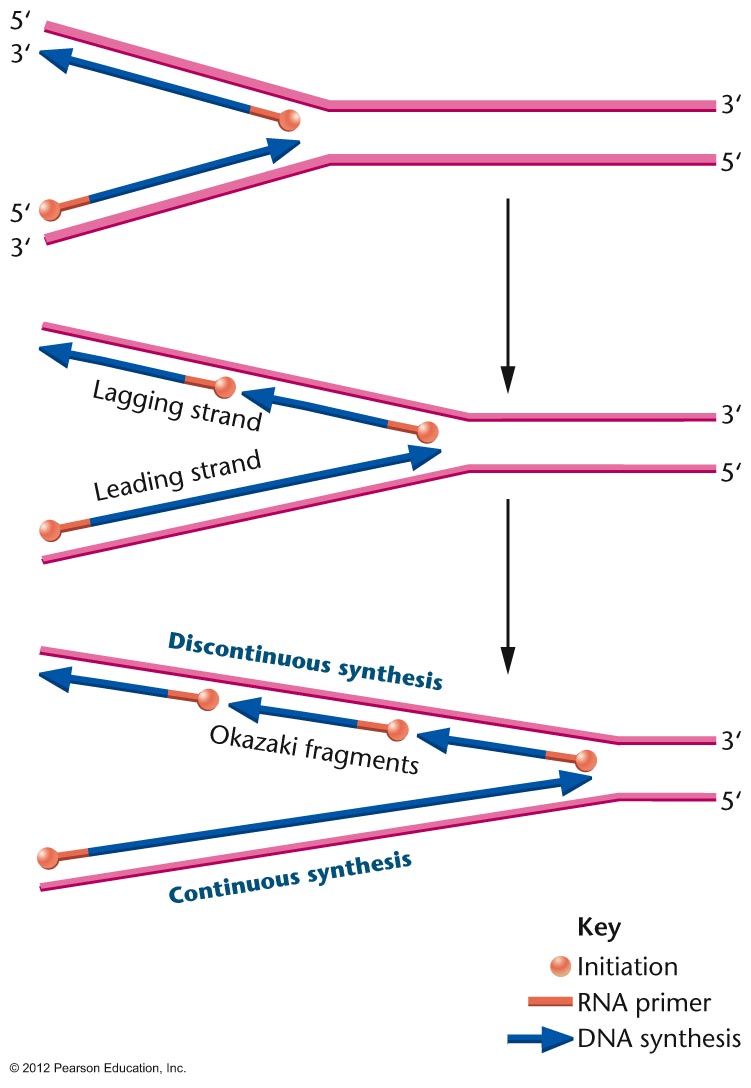
- Okazaki Fragments, Discontinuous
(Lagging Strand) and Continuous (Leading
Strand) DNA Replication:
This problem was initially solved by Okazaki
who discovered that DNA synthesis is
continuous on one strand but discontinuous on
the other. That is, he discovered that on the
discontinuous strand, first short pieces
(Okazaki fragments, 1000 - 3000 bp) are made,
then joined together. Therefore, while the
overall direction of DNA synthesis on the
discontinuous strand is 3' to 5', the actual
DNA synthesis on a molecular level is
occurring 5' to 3'. (Continuous strand
replication is also called leading strand
replication and discontinuous strand
replication is called lagging strand
replication.)
|
- 5' ---> 3' Exonuclease
Activity: An enzyme must
remove the RNA primer. This is accomplished by
some enzyme that has 5' ---> 3' exonuclease*
activity. The exonucleases involved in removing
the RNA primer removes one nucleotide at at time
from the 5' end. In E. coli, DNA
polymerase I (which has both 5' ---> 3' and
3' ---> 5' exonuclease activity in addition
to its polymerase activity) removes the RNA
primer and simultaneously synthesizes new DNA to
replace it. In eukaryotes, a special exonuclease
called RNase H appears to work along with other
exonucleases to remove the RNA primer in the 5'
to 3' directions
- DNA Ligase: The nick
left between two Okazaki fragments must be
sealed. DNA ligase joins the two Okazaki
fragments.
|
- Still Another Problem (Solved by
Eukaryotic Telomerase): The end of a
linear chromosome is called a telomere. Its
replication problem solved by an enzyme called
telomerase.
|
|
Things
I Learned at the Movies:
At least one of a pair of identical twins is born
evil.
|
 Lectures
Lectures
 Online Lectures, Quizzes, and Tutorials
Online Lectures, Quizzes, and Tutorials
{kind=link}
{kind=link}
{kind=link}
{kind=link}