|
We will now look at how
Mendel's principles can be applied more broadly. We will
also examine exceptions to Mendel's simple idea of genes,
their transmission, and their expression.
- Trihybrid
Crosses:
Mendel's same principles can be applied to crosses
involving three or more genes. In predicting the outcome
of a cross, first determine the gametes produced. In an
n-hybrid cross, the number of gametes the F1 produces is
2n, the number of F2 genotypes
is 3n, and the number
of F2 phenotypes (assuming simple dominance) is 2n.
|
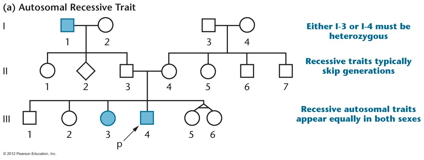
- Pedigree
Analysis:
Pedigrees are used to summarize the phenotypes of a
group of related organisms (especially humans).
|
 |
- Genetic
Notation Systems:
Two basic genetic notation systems are used.
- The "Plant System":
Gene abbreviations begin with an upper case
(dominant allele) or lower case letter.
- The "Drosophila System":
Gene abbreviations are represented by letters with
(wild-type allele) or without (mutant allele) a +
symbol. The mutant allele begins with a capital letter
if the mutant is dominant to the wild type and with a
lower case letter if the mutant is recessive to the
wild type allele (w+ is dominant to w; B is dominant
to B+).
|
|
- Exceptions to Mendel's
Idea of Dominance: There are exceptions to
Mendel's idea that when two genes are present, one is
100% expressed and the other 0%. Dominance assumes that
the expression of a gene does not depend on the number
of copies of that gene (one or two). If you look close
enough, this may not be true for any gene.
|

|
- Incomplete
Dominance
(Partial Dominance, Lack of Dominance): In
some cases, one allele is not completely dominant to
the other and the phenotype is an intermediate.
Example: Flower color in snapdragons.
|
 |
|
- Codominance:
In some cases, one allele is not completely dominant
to the other and the phenotype shows the expression of
both alleles. Examples: ABO blood group (IA,
IB) and MN group (LM,
LN).
|
- Incomplete Penetrance: This
exception occurs when a dominant allele sometimes
behaves as if it were recessive. That is, in the
heterozygote, one allele is expressed most of the
time but not always. Huntington Disease is caused by
an autosomal dominant (chromosome 4) mutation that
has about 95% penetrance. That is, about 5% of the
heterozygotes never develop the disease. (More
detail in Mutation I
outline.) Polydactyly (extra digits) of the hands is
also a dominant mutation with about 96% penetrance.
(Incomplete penetrance can be seen in qualitative
traits.)
|
- Variable
Expressivity: This exception is closely
related to incomplete penetrance but is seen with
quantitative traits. In variable expressivity, the
degree of expression (color, height, etc.) varies
among individuals with the same genotype. Variable
expressivity and incomplete penetrance have been
explained in the past as "due to differences in the
genetic background." With the advent of genomics, the
actual contributing genes in this background may be
discovered.
|
 |
- Overdominance:
In some cases, the most extreme phenotype is seen in
the heterozygote, not the homozygous dominant. Cases
of overdominance for the trait height have been
discovered in wheat and other plants.
- Sickle-Cell
Anemia: A Case Study: Sickle-cell anemia
is an inherited form of anemia caused a mutant
β-globin allele. (Normal hemoglobin has 2 β- and 2
α-globins.) Several mutations have apparently arisen
independently in malaria-prone regions of the world.
The most common mutant is due to a single nucleotide
change (A to T) of the β-globin gene, which results
in glutamic acid being substituted by valine at
position 6. This causes hemoglobin molecules to
polymerize and causes red blood cells to sickle
(elongate). These sickled cells are destroyed,
causing anemia, and also clog up organs, causing
various severe conditions. People with two copies of
the mutant allele (homozygotes) have sickle-cell
disease. People with just one copy of the allele
(heterozygotes) have sickle-cell trait. Their red
blood cells may sickle but this is much rarer, so
they have slight anemia and other problems, but not
as severely. However, in an environment where
malaria is a major health problem, people with
sickle-cell trait are actually fitter (less likely
to die). This is because the malaria parasite (a
sporozoan--which is a protist) infects red blood
cells during its multiplication stage. Cells that
are invaded by the parasite are much more likely to
sickle and therefore be destroyed--also destroying
the parasite. So, these people have much less severe
malaria attacks and are much less likely to die from
malaria. This is an example of what is called
heterozygote advantage. The term heterosis is also
used to describe cases where the heterozygote is
actually more fit than either homozygote. (Summary:
sickle-cell disease people are more likely to die of
sickle-cell disease, people with no sickle-cell
alleles are more likely to die of malaria if they
are in a malaria environment, so the fittest
are the people with sickle-cell trait.)
- Problem:
In this case, there are several phenotypes
including: 1) how sick from anemia the person is;
2) how likely the person is to experience organ
failure from sickling; 3) the person's fitness in
a malaria environment. How would you characterize
the relationship between these two alleles with
respect to these three phenotypes? (Completely
dominant, incompletely dominant, overdominant, ...
?)
- Exceptions to Mendel's
Idea of Traits/Genes/Alleles Relationship:
Mendel proposed that for each trait there was 1) one
gene consisting of 2) just two alleles. There are
exceptions to both of these postulates.
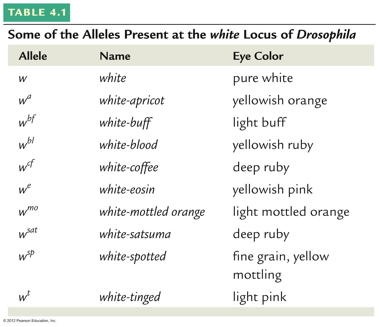
- Multiple
Alleles: Some genes (actually all genes) have
more than just two alleles, although each individual
has only two. Example: ABO Blood Group (IA,
IB, i). Although this is a complication of
Mendel's ideas, it does not alter how you would
predict outcomes of crosses.
- Multiple
Genes, Polygenes, Quantitative Traits,
Mutiple-Factor Traits, Multifactorial Traits:
Some traits are determined by more than one gene.
(Theoretical) Example: Plant height (determined by two
or three genes with incomplete dominance). These are
usually continuous (vs. discontinuous) traits.
(Continuous traits=quantitative traits; discontinuous
traits=qualitative traits). With environmental
influence and variable expressivity, these traits can
truly become continuous. The color of wheat grain is
determined by a polygene (3 genes, 7 phenotypes,
1:6:15:20:15:6:1 ratio in the F2. Quantitative traits
(including human intelligence) are determined by
heredity and environment. The quotient called
heritability (h2) measure the proportion genetics
contributes to a trait (h2 = 1.0 means the trait is
completely determined by genetics; h2=0 means the
trait is completely determined by the environment).
(Human multifactorial traits, twin studies, and DNA
methylation: intelligence,
psoriasis,
Genetics
Society of America statement on race and
intelligence--scroll down to 1975.)(Equally additive
genes problem)
- Human
complex multifactorial traits: Many human
traits (other than intelligence) are multifactorial
traits and also have an environmental component.
- Autism
has been shown to be influenced by a number of
genes (AUTS1, AUTS3, AUTS4, and AUTS6 through
AUTS17, all of which are autosomal, in addition to
4 X-linked susceptibility loci).
- New Research
on Educational Achievement:
The General Certificate of Secondary
Education (GCSE) exam is administered at the end
of compulsory education at age 16 in Great
Britain. GCSE scores were obtained for 13,306
twins at age 16 and assessed on 83 scales that
were condensed to nine broad psychological
domains, including intelligence, self-efficacy,
personality, well-being, and behavior problems.
The mean of GCSE core subjects (English,
mathematics, science) is more heritable (62%) than
the nine predictor domains (35-58%). Each of the
domains correlates significantly with GCSE
results, and these correlations are largely
mediated genetically. The main finding is that,
although intelligence accounts for more of the
heritability of GCSE than any other single domain,
the other domains collectively account for about
as much GCSE heritability as intelligence.
Together with intelligence, these domains account
for 75% of the heritability of GCSE. We conclude
that the high heritability of educational
achievement reflects many genetically influenced
traits, not just intelligence. (PNAS,
October 21, 2014)
|
|
- Epistasis:
This type of gene interaction occurs when one gene
masks the effect of another. The gene that is doing
the masking is the epistatic gene and the one that
is being masked is the hypostatic gene. Example:
Albinism is an example of recessive epistasis (C_ is
pigmented, cc is albino; B_ is agouti, bb is
black--note cc is albino regardless of which B/b
genes are present). Example cross: agouti x white
(P) ---> agouti (F1) ---> 9 agouti : 3 black :
4 white. Albinism is often a mutation in the
tyrosinase gene. Tyrosinase catalyzes an initial
step in the synthesis of melanin (tyrosine
--> ).
- Gene
Suppression: A special type of epistasis
is gene suppression where a second mutation
reverses the effect of a first mutation. Example:
Drosophila:
f+/_ females
have normal bristles and f/f females have short,
bent bristles but f/f su(f)/su(f) females have
normal bristles.
|
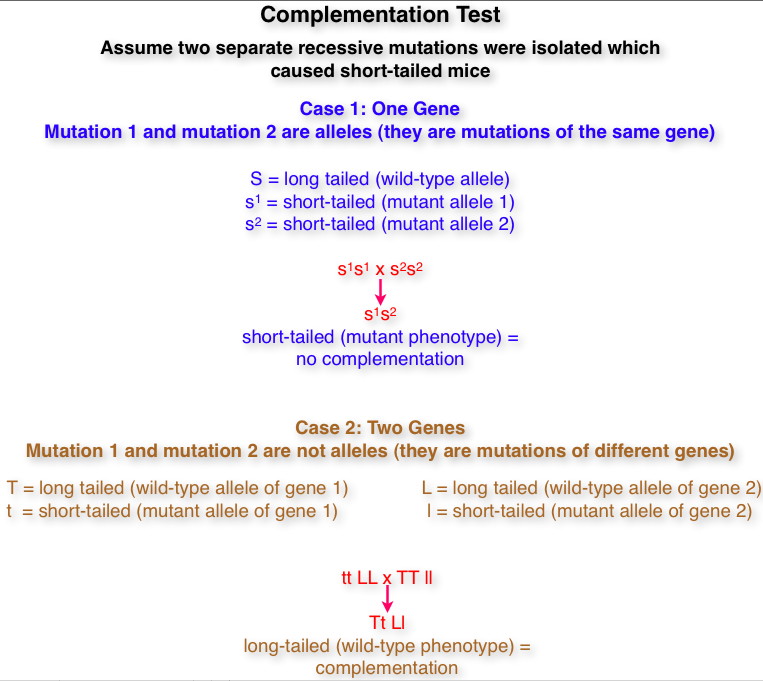
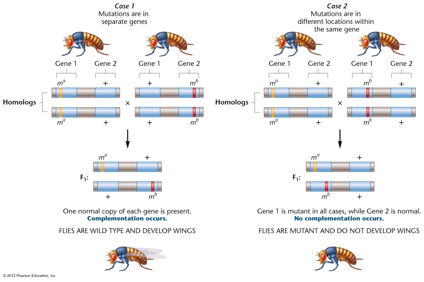
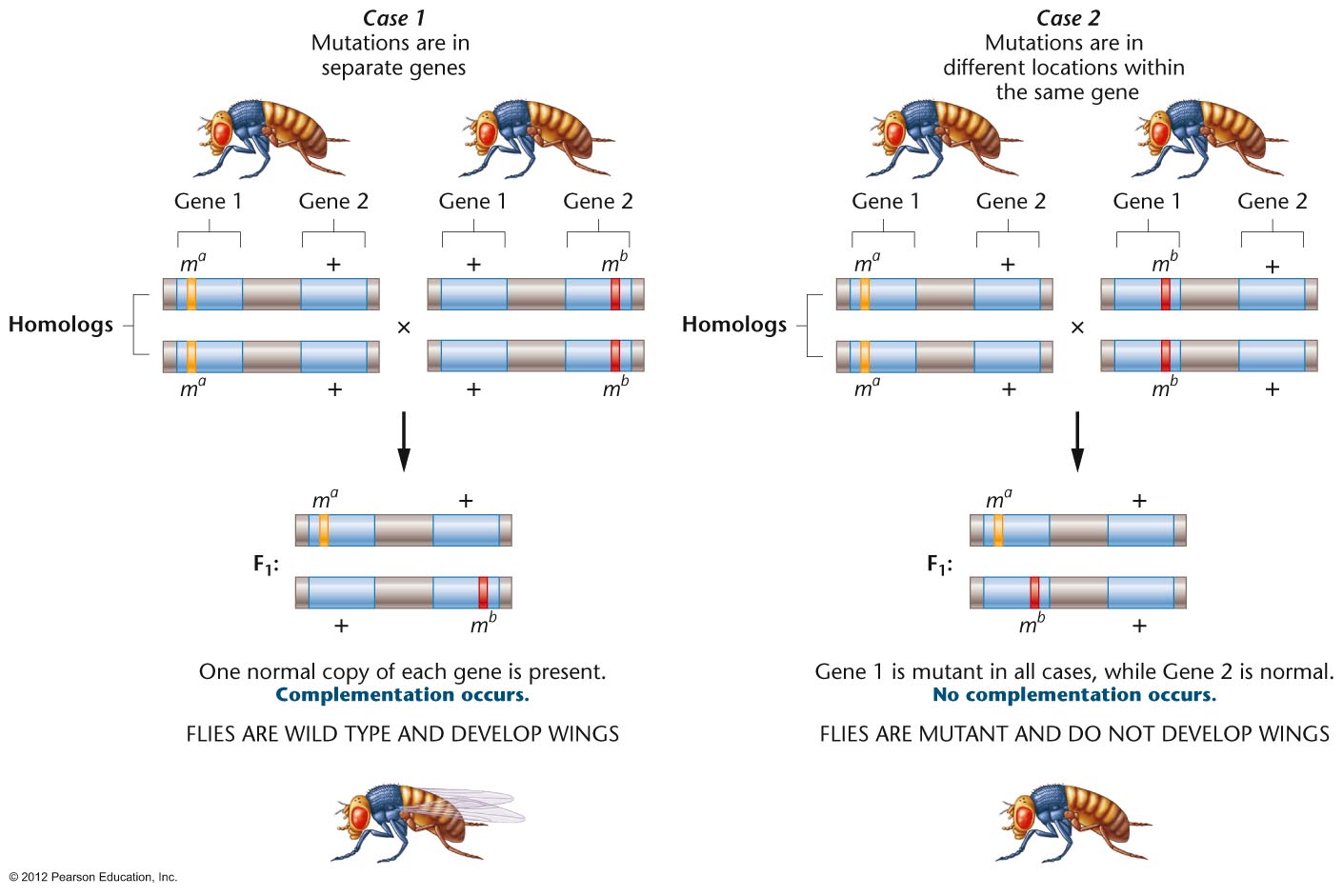
- Complementation Test:
Since a trait may be determined by one gene or two or
more genes, how do we know if two mutations are alleles
(the same gene) or not alleles (different genes)? A
complementation test is performed (cross homozygotes for
the two mutations). If the offspring are wild type,
that is if the genes complement each other, then they
are not alleles. If the offspring are mutant, that is if
they do not complement each other, they are alleles.
(There are some rare exceptions to this rule where
alleles will complement each other: intragenic
complementation or interallelic complementation).
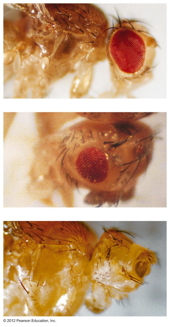
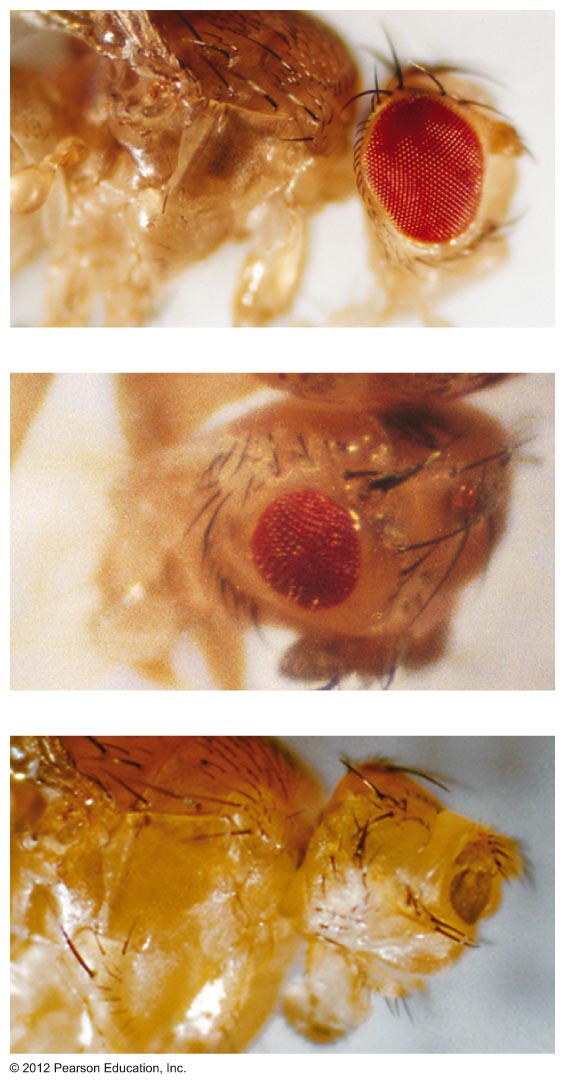
- Pleiotropy: In this
exception to Mendel's idea of the relationship between
traits and genes, one gene can effect more than one
trait. There are numerous examples of pleiotropy in all
organisms (white eye, w
in Drosophila,
yellow in mice).
|
|
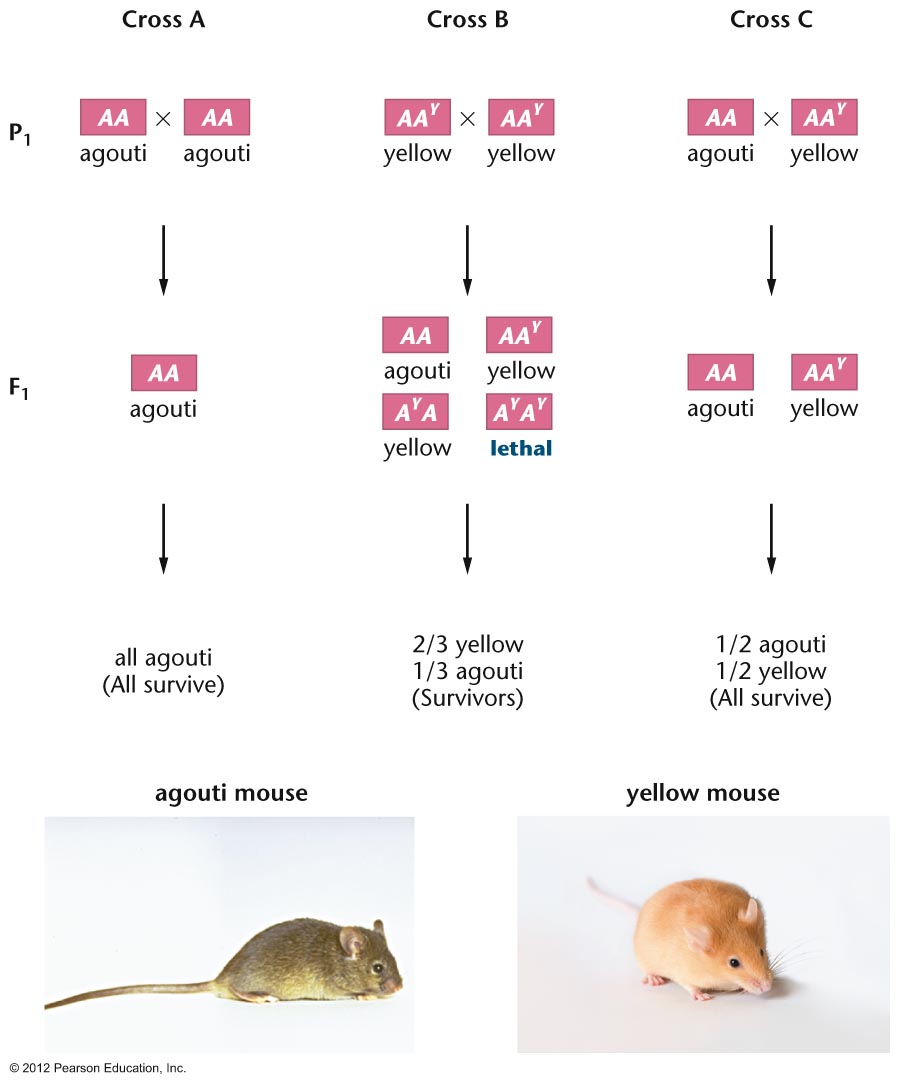
- Lethal Genes: Genes
that cause death can obviously alter Mendelian ratios.
There are recessive lethals (yellow in mice) and
dominant lethals (Huntington Disease), although usually
dominant lethals do not stay around long. When two
yellow mice are crossed, the offspring are: 1 agouti : 2
yellow (yy is agouti, Yy is yellow, YY dies).
|
|
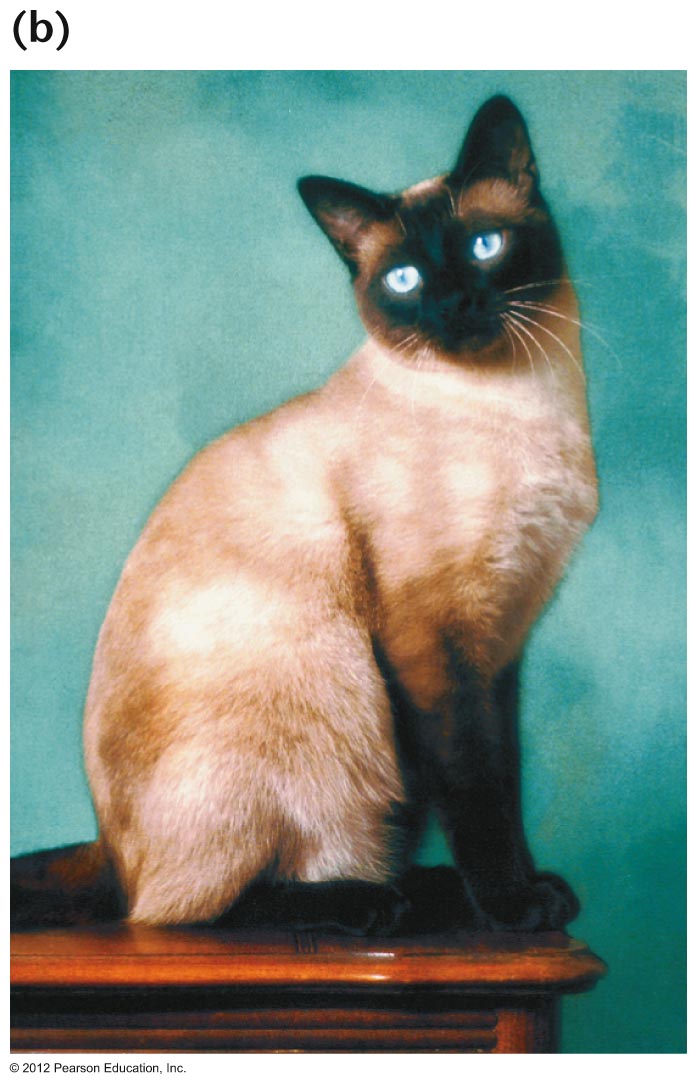
- Conditional
Mutations/Temperature Sensitive Mutations: The
expression of some genotypes is dependent upon
environmental conditions, like the temperature at which
the organisms is reared. Siamese cat coat color is
an example. It is due to a temperature sensitive
mutation in the tyrosinase gene such that homozygotes
have white fur where body temperature is high and black
where it is low. Temperature sensitive mutation and
conditional lethals have been valuable in studying gene
function.
|
|
- Position Effect:
The expression of a gene can be altered by moving it to
a new chromosomal location (to a new genetic
neighborhood). These effects may be related to
heterochromatinization or other effects.
- Maternal Effect:
The phenotype may be influence by the genotype of the
mother (the egg). One example is the direction of
coiling in shells of the snail Limnaea, as
described in a paper
by Morgan's student Sturtevant in 1924. Right-handed
coiling is the result of the dominant allele L
while left-handed coiling is caused by its recessive
allele l. However the direction of coiling of
the snail's shell is determined by not the genotype of
the snail itself, but by its mother's genotype. Therefore,
an LL or Ll snail will have all
right-handed
- Linkage and Sex Linkage:
The most significant exception to Mendel's ideas came
with the discovery that genes are on chromosomes. We
will take up linkage (non-independent segregation) in
the next unit.
|
 Lectures
Lectures
 Online Lectures, Quizzes, and Tutorials
Online Lectures, Quizzes, and Tutorials



{kind=link}
{kind=link}
{kind=link}
{kind=link}